
Man has been a part of the animal kingdom for more than three million years. Yet many cultures have seen man as quite separate, or excluded man entirely, from nature. We see this among monotheistic societies that nurture theological explanations for the origin of our species. Prior to the Scientific Revolution, theological considerations served largely to constrain inquiries in this area. The naturalist movement that emerged during this period enabled some finally to break free of scriptural assumptions.
Jean-Baptiste Lamarck held the chair in invertebrate zoology at the Muséum national d’Histoire naturelle (MNHN) in Paris, when he published Recherches sur l’organisation des corps vivants (Research on the Organization of Living Bodies) in 1802. This small, but revolutionary, book contained the first theory of evolution applied to the origins of Homo sapiens. In this view the progressive straightening of the vertebral column seen in the first hominids was driven by climatic changes that forced some apes to adopt walking over arborealism. This idea was then popularized by Charles Darwin in The Descent of Man, published in 1871. It is now the consensus in paleoanthropology.
A very different reality is reflected in the collections of the MNHN, on which I have based my work. On the contrary, straightening, with its correlated anatomical and psychomotor changes, is an intrauterine process that took place over the course of more than forty million years. It resulted from a growth in complexity of the embryonic nervous system and its rotational dynamics, and led to a succession of threshold effects incompatible with the nested hierarchy of Linnaean classification.
The identification of the first hominids is flawed.
Linnaean Classification
Lamarck was a disciple of the eminent naturalist Georges-Louis Leclerc, Comte de Buffon. From 1730 until his death in 1788, Buffon was the director of the Jardin du Roi in Paris.1 Known today as the Jardin des Plantes, it encompassed not only a vast herbarium, but also the Cabinet d’histoire naturelle. This was, at the time, the largest such collection in Europe. It was opened to the public by Buffon in 1739.
Naturalists of the era, curious about the structure of living organisms and the manner in which this structure is passed on from one generation to the next, tended to follow the philosophical tradition of Gottfried Leibniz. In his posthumously published Protogaea, Leibniz speculated about the formation of the earth, and suggested that fossils were the remnants of animals “qu’on ne trouve pas aujourd’hui.”2 Leibniz also advanced the principle of natura non facit saltum: nature does not make jumps.
Carl Linnaeus, a contemporary of Buffon, introduced a taxonomic hierarchy in Systema Naturæ (1735) and in his later works. Organisms belong to a single species if they produce generations of offspring that can interbreed. Conversely, two organisms with similar anatomies, but unable to interbreed, form two different species that fall under a second category, the genus. Similar genera are classed under a third category, the family, which is in turn classified under order, order under class, and class under kingdom. A nested system of classification emerges from this structure, one that is entirely a product of the human mind.
According to Linnaeus, man is a species, sapiens. Anatomical differences between sapiens and the species that were considered closest at the time—the Angolan chimpanzee, Bornean orangutan, Southeastern Asian gibbon, macaque, and other African and American monkeys—sufficed to distinguish at least two genera: Homo and Simia. Sapiens was assigned to Homo, the others to Simia.3 The anatomical definition of the genus Homo was thus conflated with the species sapiens. The definition of a species, it should be noted, refers to an objectively real phenomenon. Simia were also close to a third genus, Prosimia, comprised of small arboreal mammals such as the Madagascar lemur and the lorises and tarsiers of southeast Asia. These three genera are classified under the order Primates.
Buffon, however, rejected Linnaean classification as an objective representation of the animal kingdom, claiming that “in nature there really exists only individuals; genera, orders, and classes exist only in our imagination.”4 The notion that Homo sapiens should be classified as a primate was also dismissed by Buffon. Primates, he pointed out, do not possess a unique anatomical organization, of which Prosimia, Simia, and Homo sapiens are simply variants.5 No element of the morphological organization of Homo sapiens can be confused with that of a Prosimian. In the case of primates, in fact, Linnaean classification groups together several anatomical configurations without taking into account their more extreme variations. It is not a suite of species that diverges, by small gradations, from lemur towards Homo sapiens.
No primatologist working today would consider Homo sapiens an augmented prosimian, which defines “primate morphology.” And yet they would also place Homo sapiens within the order of primates. Morphological discontinuity is a fact.
Why this paradox?
Lamarck’s Revolution
Lamarck took charge of the botanical collections at the Jardin du Roi in 1789. Following the Revolution, Lamarck became deputy to the démonstrateur des collections, Louis-Jean-Marie Daubenton, and assisted Buffon in reorganizing the Cabinet du Roi. In 1793, when the Jardin du Roi was reorganized into the MNHN, Lamarck held the first chair in invertebrate zoology.
In 1764, based on a study of the skulls of dogs, lemurs, chimpanzees, and Homo sapiens in the MNHN collection, Daubenton made the first external interspecific comparison of the foramen magnum, the hole at the base of the cranium through which the spinal cord enters the skull. The foramen magnum is oriented backwards in dogs and Prosimia, downwards and forwards in Homo sapiens, and is intermediate in Simia.6
Figure 1.
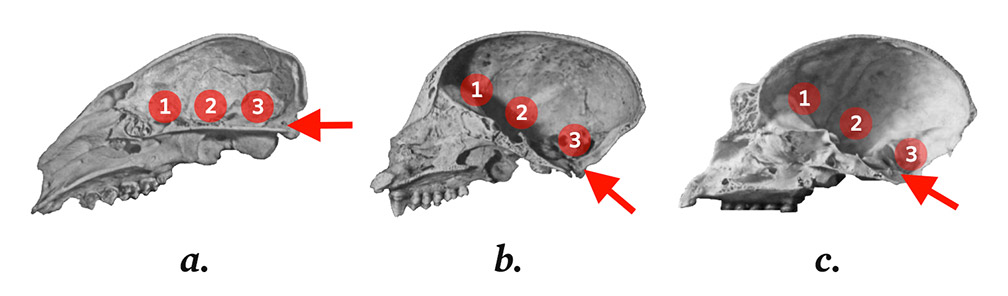
Clockwise from top left: (a) Prosimia (lemur) without basi-cranial flexion; (b) Simia (American monkey) and (c) small Asian anthropoid (gibbon), both with the same flexion; (d) African great ape (chimpanzee) with greater flexion; and (e) Homo sapiens, with a verticalized flexion.7 The arrow indicates the foramen magnum, and the two broken lines inside the skull form the sphenoidal angle.8
Lamarck was familiar with Daubenton’s work; after all, they had collaborated for a decade. In 1802, two years after Daubenton’s death, Lamarck published Recherches.
In order to change any interior system of organization, a combination of circumstances more influential and of much longer duration is necessary than that required to alter and modify the external organs. But I have observed that when circumstances require, nature moves from one system to another, without making a jump, if they are neighbors. Indeed, it is by this faculty that she has managed to form them all successively, by proceeding from the simple to the more compound.9
Lamarck extrapolated this idea to the origins of Homo sapiens, even though no primate fossil had yet been found. Lamarck proposed a thought experiment:
As for man, the closest to the quadrumanes [a classification of primates with four hands, such as the chimpanzee] is strongly distinguished by a great superiority of intelligence and also very much by numerous traits of organization that characterize him: 1) the foramen magnum, which is set completely within the base of the skull, so that the head is at the top of the vertebral column and he can see a much greater number of objects at once; 2) the remarkable mobility of his fingers; and 3) the fact that, by the state of his organization, man is made to stand and walk upright. Can we not think that this state of organization has been acquired little by little over a long period of time, with the help of circumstances that became favorable?10
The ever-expanding fossil collection at the MNHN led to the creation of two new disciplines, paleontology and comparative anatomy. Their founder was Georges Cuvier, who held the first chair in comparative anatomy at the MNHN, a position created in 1802.
Cuvier developed a systematic approach to anatomical organization in which function shapes form.11 He went beyond Lamarck by stressing the principle of the correlation of parts. If a piece is missing, assembly of the whole is no longer possible. This conflicts with Lamarck’s idea of gradual transformation, itself founded on another of Leibniz’s postulates: a living structure cannot be reorganized both simultaneously and globally.
In accordance with Buffon, Cuvier accepted the idea of the extinction of a species, and considered climatic crises to be the primary factors. It was inconceivable to Cuvier that changes occurred through small graduations over long periods of time; if one part changed, the whole living system needed to be reorganized. Prosimia, Simia, and Homo are not variants of a single primate anatomical system or germ line. Cuvier and his disciples could not move beyond their shared conviction that nature does not make jumps.
This was not the only difficulty. Microscopic examination revealed a particular chronology in structural development, even at the level of a seed from the time of its fertilization. There was nothing visible that might correspond to a form of memory, or what we might now call heritability, shaping the living organization of a species. Neither Lamarck nor Cuvier could have encountered underlying physical or chemical features that might have led them to imagine a causal link between a change in habitat and the coordinated evolution of an organism in its entirety.
Cuvier and his followers faced profound difficulties in accounting for the morphological organization common to the chimpanzee and the orangutan as opposed to that of Homo sapiens.
Man in Nature
Paul Broca, a professor in the faculty of medicine at the University of Paris, founded the Société d’Anthropologie de Paris (SAP) in 1859.12 Two years earlier he had founded the first public center for teaching anthropology. Broca was among the leaders of European physical anthropology. Among Broca’s many contributions was his work in anthropometry, the science of studying the measurements and proportions of the human body, and, in particular, in craniometry. Progress in this field meant that forms of morphological organization in which both constants and variables are present could be described quantitatively. It now became possible to verify whether the variation among primates was of a structural nature, or, as Buffon and Cuvier believed, the structure common to individuals is precisely that which does not lend itself to variation.
Five decades after the publication of Recherches, the Lamarckian revolution continued to exert an influence, especially on Darwin, who published On the Origin of Species in 1859. Still not in question, however, were the origins of Homo sapiens. Thomas Henry Huxley’s The Place of Man in Nature, published in 1863, placed Homo sapiens within the order of primates.13 Five years later, the presentation of a French translation of The Place of Man in Nature to the members of the SAP triggered a series of debates on evolution. The discussions set Broca against Armand de Quatrefages, who held the chair in anthropology and ethnography at the MNHN, and was a disciple of Cuvier and member of the Academy of Sciences. De Quatrefages believed Homo sapiens was itself an order, bimana (apes with two hands and two feet), and not a genus within the order of primates. For Broca, transformism remained a delicate and potentially controversial question which should not interfere in questions of classification. Broca openly addressed the issue at the first meeting of SAP in 1869, asking that the group stick to observation:
The question that M. [Eugène] Dally [translator of The Place of Man in Nature] has promised to treat was a question of pure zoology. It was a matter of knowing if one should keep the classical distinction of the order of bimanes and the order of quadrumanes. … Today I will merely consider what place comparative anatomy should assign to the genus Man in the classification of mammals. The characteristics that distinguish this genus from others—do they have the weight of those that constitute classes, or subclasses, or orders, or only families?14
Broca subsequently published a study presenting the positive aspects of Linnaean classification, as well as a theory of the natural and increasingly close connection between anatomical organizations from Prosimia to Homo sapiens.15 It was in 1858 that Rudolf Virchow defined an angle in the skull base, the sphenoidal angle, that would allow quantitative measurement of the straightening of the nervous system. Yet differences in Virchow’s sphenoidal angle alone, Broca pointed out, demonstrate that Homo sapiens will never be counted among the varieties of great apes. Rather, they justify distinguishing Homo sapiens as a family apart from the chimpanzee and orangutan.
Broca distanced himself from de Quatrefages, but he also insisted on maintaining a distinction between Homo sapiens and apes, in order to elevate human anatomy to the rank of a family, the Hominidae:
Man evidently differs from apes more than two species of the same genus differ, more than two genera of the same family differ … Man cannot constitute more than an order, or less than a family.16
In 1871, Darwin published The Descent of Man. “The conclusion that man is the co-descendent with other species of some ancient, lower, and extinct form, is in not in any way new,” Darwin asserted, noting that, “Lamarck long ago came to this conclusion.”17 Darwin was neither an anthropologist, nor a primatologist. Nonetheless, he adopted Lamarck’s hypothesis for the origins of man.
As soon as some ancient member in the great series of the Primates came, owing to a change in its manner of procuring subsistence, or to a change in the conditions of its native country, to live somewhat less on trees and more on the ground, its manner of progression would have been modified, and in this case it would have had to become either more strictly quadrupedal or bipedal.18
Darwin and his contemporaries were themselves part of the Lamarckian revolution. A function gives rise to an organ and then the organ, including the shape of its bone tissues, is gradually modified. Those changes are then, through use, transmitted to later generations.
Darwin did not invent the idea of a common ancestor for the chimpanzee and Homo sapiens, nor that evolution was a result of behavior, nor that modifications adapted by choice are conserved. The concept of natural selection is merely a formulation of the Lamarckian idea of the conservation and transmission of modifications, constrained by climatic changes. It arose from the gradualist model and was only applicable to small variations that would not affect the overall organization.
Darwin’s essay on the origins of the genus Homo was a thought experiment, a simple idea projected into the past.
The Princeton Doctrine
Darwin was constrained by a lack of the kind of biophysical or biochemical knowledge that would have allowed him to judge the limits imposed on the inheritance of acquired characteristics. In 1927, Hermann Muller highlighted the close relationship between the molecular composition of the cell nucleus and heredity. He irradiated Drosophila larvae with X-rays, accelerating their mutation rate and producing deformed flies. Whence the idea that large molecules could function as a form of biological memory.19
In January 1947, George Simpson, Ernst Mayr, and Theodosius Dobzhansky organized a conference at Princeton University entitled “Genetics, Paleontology and Evolution.” A new synthetic theory of evolution was developed. The Princeton conference marked a radical break with the Lamarckian–Darwinian school, which had focused on the need for organisms to adapt to ecosystems. This was no longer the case for the modern synthesis. The authors developed their doctrine by focusing on random genetic mutation, each random change representing a kind of memory loss. Organisms were no longer regarded as responding to environmental change, but as resulting from random mutations.20 Under neo-Darwinism, orthogenesis disappears, the concept of a body plan shattered.
In April 1947, Jean Piveteau, Camille Arambourg, and Pierre-Paul Grassé organized a famous international symposium devoted to paleontology and evolution. They invited the most significant figures in evolutionary biology. Piveteau wrote:
Those which the paleobiologist notices and wishes to explain are the large evolutionary changes which make not only simple quantitative modifications, but qualitative transformations, leading to true, organic redesigns. Is there not, in moving from small divisions to large divisions of classification, a necessary appeal to explicitly new factors? One of the problems which it is essential to expose precisely is that of the relations between microevolution and macroevolution.21
Marcel Prenant described the Lamarckian school, which attributes a designer-like role to the environment, as barely rational.22 For the paleontologist David Watson, distinctions among large systems of organization did not require further investigation.23 Simpson agreed.24 But Arambourg demurred.
One cannot, in fact, consider orthogenesis as an internal tendency of mysterious nature. But we are nevertheless obliged to note that since the origin of life, one finds a closer and closer adaptation of organisms to various biotopes … It is certain that one finds, without seeing precisely the cause, increasingly complicated organisms.25
He noted that fossil specimens had evolved in the same direction with changes in the humidity of the environment, and did so far too rapidly for genetic accidents to have modified the adaptations in the direction evident from the fossils.
The facts of paleontology were primary; they had to be understood.
Arambourg was not a neo-Darwinian. He was a punctualist, a saltationist, and a Lamarckian:
The well-known facts of morphological convergence in groups with very diverse origins, but living in identical environments, seems to me to be difficult to explain, other than by a certain action of the environment, that is to say by the intervention of a “Lamarckian” cause.26
Population genetics did not explain macroevolution, a difficulty since the time of Buffon and Cuvier. With his usual skill, J. B. S. Haldane summarized the situation:
In the field of genetics, I think everyone who has tested it, admits Mendelian inheritance of certain characteristics at least. … Extranuclear heredity exists, above all in green plants … L’Heritier has proven that the sole characteristic of extranuclear heredity discovered in Drosophila melanogaster can be transmitted by grafting, and its agent can be considered like a virus.27
New Hominids
In April 1947, human paleontology took a fresh twist with the discovery of the most complete australopithecine skull found to that date. The skull, informally known as Mrs. Ples, was found by Robert Broom and John Robinson near Sterkfontein in South Africa.28 It is small, roughly the size of a chimpanzee’s skull, obviously non-human and prognathous, the foramen magnum oriented to the front and downward, and the Virchow angle is closed. Other fragmentary finds at the site, possibly from the same individual, included a short, broad pelvis that does not resemble that of any known ape, and the head of a femur adapted for permanent bipedalism. Together these fossils showed a transformed skeleton, Australopithecus africanus, verticalized from the center of the skull base to the sacrum.
Additional discoveries during the 1970s, such as the finds in the ancient lakeside and fluvial deposits of the Great Rift Valley in Ethiopia, had a lasting effect on the field. Up until then, paleontologists had been creating another group with extinct genera, Archanthropians, to classify intermediates between the great apes and Homo sapiens: Pithecanthropus by Eugène Dubois in Java, Sinanthropus by Davidson Black in China, Atlanthropus by Arambourg in Algeria, and Telanthropus by Broom and Robinson in South Africa. The Paleoanthropian group was created for Neanderthal man.
On the Linnaean system of classification, it should be possible to distinguish a structure that elevates Homo sapiens to the rank of genus when compared to the characteristics of the species. Homo anatomy must appear gradually before any sapiens characteristics. For obvious scientific reasons the genus Homo must be rigorously defined. The defining characteristics of the skeleton of Homo sapiens are, in fact, linked to the degree of verticality in the brainstem and spinal cord. Homo corresponds to a verticalization now visible only in sapiens.
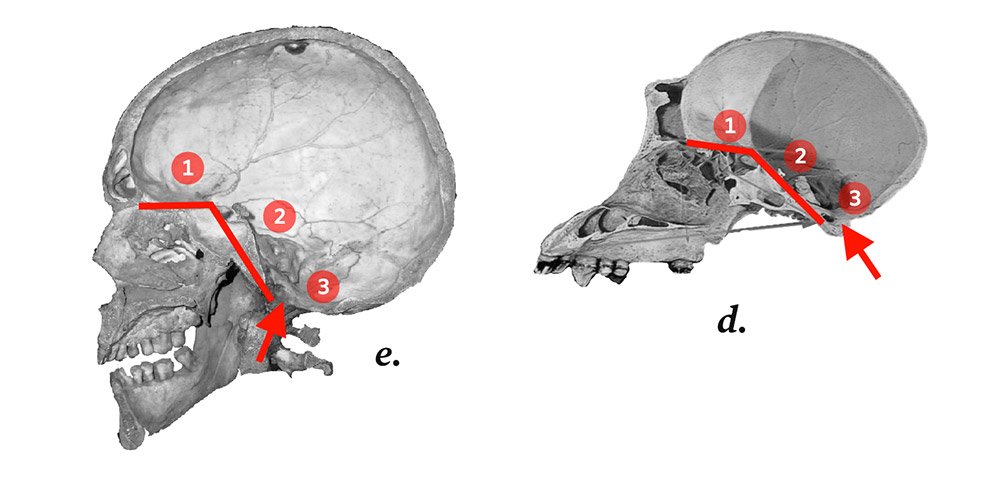
In 1947, Arthur Keith argued that a brain volume of 700–800cm3 marked the so-called cerebral rubicon between apes and men.29 This figure was widely accepted until 1964, when Louis Leakey et al. published a paper describing a skull discovered at Olduvai Gorge in Tanzania.30 They proposed downgrading the cerebral rubicon to a brain volume threshold of 600cm3, and called their skull Homo habilis. As well as adjusting the boundary at which a fossil species defined the genus Homo, this approach ensured that their newly discovered species, habilis, could be accommodated within the genus.31 None of the habilis cranial bones, from mandible to neurocranium, corresponds to verticality or the morphogenesis of Homo sapiens. The habilis fossil marked a genus, but not one that could be used to define the degree of upright posture around which our anatomy is organized.
Figure 2.

Clockwise from top left: Comparison of the basicranial straightening (shown in red, with 1 and 2 marking the angle) in (a) Gorilla, (b) Australopithecus africanus, (c) Homo habilis, (d) Homo sapiens, with (e) a drawing of the sagittal section of the human brain.32
In 1974, Yves Coppens codirected an international mission in Ethiopia that discovered the near complete australopithecine skeleton known as Lucy. Eight years later, he presented the East Side Story, a theory that attributed the origin of hominids to the collapse of the East African Rift and a resulting imbalance in rainfall. Continental rainfall west of the Rift would have preserved the subtropical forests, while the forest would have been depleted within the less well-irrigated Rift. Apes that remained in the Rift would have been forced to begin moving around on two legs.33 The theory proved popular until doubts arose from studies of the australopithecine musculoskeletal system. The australopithecine pelvis and ankles were adapted to a vertical posture, but australopithecines were also arboreal, as shown by the joints of the limbs, shoulders, and fingers. They were no less arboreal, in fact, than the chimpanzees that currently inhabit the Victoria and Tanganyika lake regions, likely the descendants of apes that inhabited the area between twenty and twenty-three million years ago.
At the end of the twentieth century, the theory that the origins of upright posture originated in climate change was abandoned by Phillip Tobias, a co-author of Leakey’s famous 1964 paper. East and South African habitats, Tobias argued, were a mosaic, ranging from gallery forests to woodlands and savannahs. Ten years later, paleontologists exploring the Ethiopian Rift recognized that Australopithecus afarensis and their quadruped arboreal predecessors, Ardipithecus, also lived in a mosaic environment.
If so, what was the engine driving the development of upright posture?
Mandibles and Skull Bases
My own research suggested that changes in mandibular shape, during ontogeny and phylogeny, reflect the straightening of the skull base and mark the inception of upright posture. These changes had nothing to do with diet and chewing, or locomotion. The mandibular joints articulate with the erect part of the skull base and depend on both the neural straightening and the transversal development of the cerebral hemispheres. The mandible and the degree of verticality of the skull base are both sapiens during embryogenesis. Mosaic evolution did not apply to the cranio-caudal body plan of primates. Neural trajectories change during embryo formation; neural straightening results as successive thresholds are crossed. This straightening, and thus the vertical positioning of the cerebellum, takes place in the amniotic fluid at scales of a few millimeters, along with a complete reorganization of the spatial morphogenesis of the supporting tissues that will form the skeleton.
Comparing the morphology of the mandible among fossil and living great apes, Homo sapiens, and fossil hominids revealed a dynamic link between the upright posture of the chin and the cervical column. My early work was focused on the morphogenesis of the mandible from the period of intrauterine growth through to adulthood. Measurement protocols were defined and applied to the widest possible selection of primate species. In accordance with this classifications, six morphogenetic groups have been identified:
- prosimians, including Tarsius;
- simians, monkeys, and gibbons;
- apes;
- Australopithecus (and its close relative, Paranthropus)
- all the species previously classified as Archanthropians and Paranthropians: habilis, ergaster, rudolfensis, erectus, and neanderthalensis; and
- Homo sapiens.34
Figure 3.
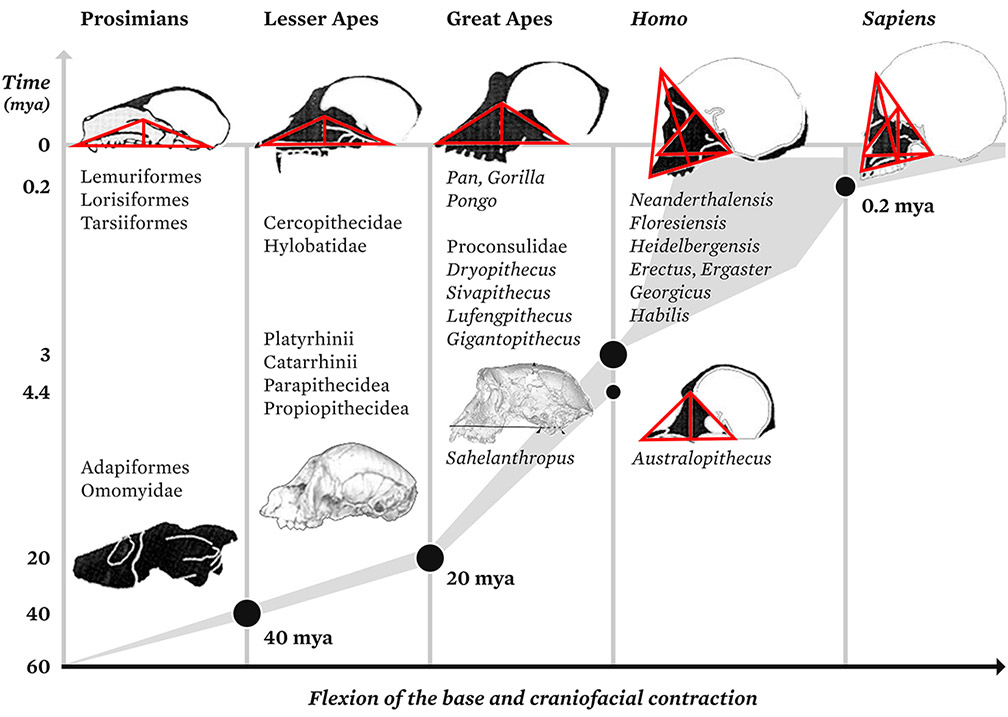
Phylogenesis of embryonic straightening in the adult skull base. (mya = million years ago)
It soon became apparent that the Virchow angle was predominant in determining the general architecture of the skull base and face during intrauterine development.35 In 1924, Louis Bolk proposed a theory of fetalization. From his observations of mammalian embryos, Bolk inferred that the posterior base of the cranium was verticalized from the very first stages of embryogenesis. In small-brained species, the Virchow sphenoidal angle declines over the course of fetal development and, by birth, the base of the skull has become flat. The vertebral column appears as it lengthens and is found behind the skull in all mammals except simians and, in particular, Homo sapiens.
Bolk proposed that Homo sapiens retained an ancestral form of Virchow folding due to the longer fetal growth period of the cerebral hemispheres whose trajectory rotates around an axis that passes through the center of the skull base.36 During this extended period of growth, rotational pressure is applied to the folding until birth. This results in the upright posture of the posterior base. The neurocranium develops for a longer period, to the detriment of the face. Facial development is delayed and then halted following birth, which reduces prognathism.
The embryogenesis of the head involves the development of tissue situated above the skull base that becomes the brain, cerebral trunk and cerebellum. This tissue is the cephalic extremity of the embryonic neural tube, from which the brain and spinal cord form. The rotation of the cephalic extremity, upwards and toward the back, is greater in amplitude in Homo sapiens than in a mammal without flexion. The upright posture of the skull base and face, or cranio-facial contraction, is therefore linked to the duration of neurogenesis. From the prosimians to Homo sapiens, phylogenesis was a succession of six neurogenetic stages.
In 1900, G. Levi published photographs of exceptional quality showing the different stages of embryogenesis of the base of the skull.37 At stage nineteen, between the sixth and the seventh weeks, a contraction can be observed, forming a step like a closing accordion. At the end of the eighth week, the flat part of the step straightens out. The profile can be measured by reference to Virchow’s sphenoidal angle. Levi described the cartilaginous base, abstracted from its anatomic context, but did not offer an explanation for these changes. I conjectured that morphogenesis depended on internal constraints at an infra-millimeter scale. Because the trajectory described a rotation, it was necessary to define an appropriate reference frame. I chose the dorsal cord that first appeared six hundred million years ago at the base of the vertebrates, and that extends under the neural tube. The cephalic termination of the dorsal cord defines an abscissa and an ordinate.
A dynamic picture emerges showing the movement of the neural tube up until the stage where the steps first contract, then a second movement which straightens out the posterior segment of the base. This synthesis demonstrated for the first time the origin of the verticalization of the nervous system, the skull base, and the resulting mandibular changes.38
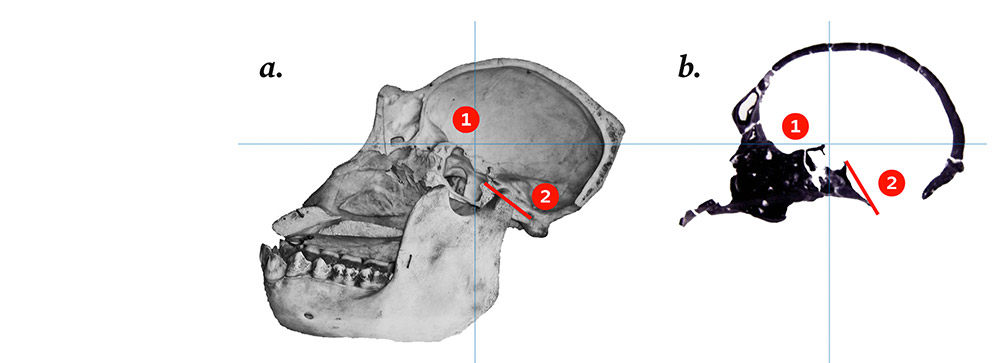
Figure 4.

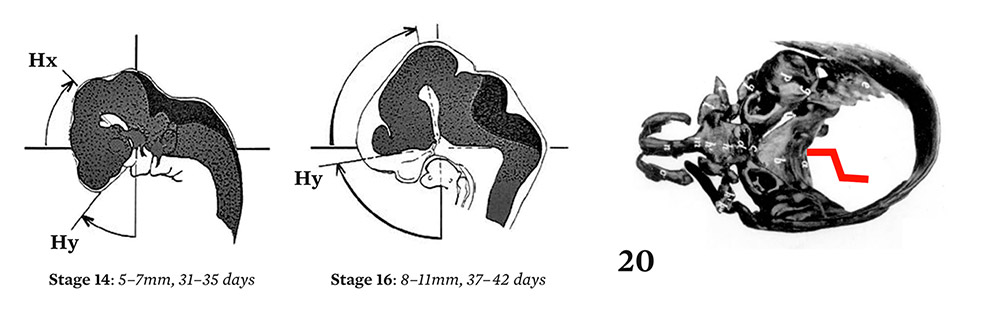
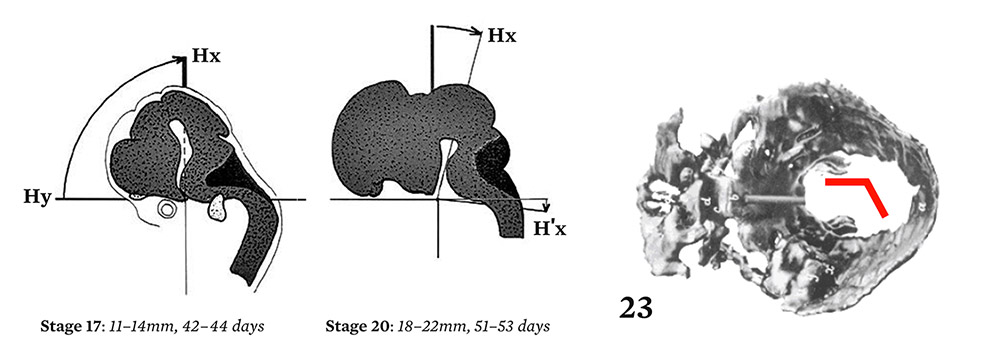
Elongation and rotation of the human neural tube with reference to the dorsal cord, showing dynamic links with the folding of the chondrocranium between stages 20 and 23.
Confirmation for this thesis emerged from an animated study of microscanned human embryos.39 A distinctive rhythm of expansion and contraction is evident in the neural tube throughout the animation. The embryos of tarsiers have been studied using the same techniques, providing a view of the tissues in median sagittal section. Comparisons with homologous stages of Homo sapiens provided spectacular confirmation that during stage nineteen something happened each time the nervous system straightened.
These embryological details have dramatic consequences. The emergence of upright hominid posture need no longer be linked to habitat changes. Its origin must be attributed to the increasing complexity of the nervous system. The embryonic body plan was reorganized through a series of threshold effects which are still in evidence in every human embryo.40
Paleoanthropologists working in this area have focused on the semicircular canals that allow the cerebellum to control the position of the head and of the body. The canals are in the petrous bone, near the center of rotation. This has long been interpreted as the result of large-scale base folding. A recent discovery has overturned this idea. The canals of Australopithecus remained simian while its cartilaginous base was verticalized. It is not precisely the process seen later in Homo habilis. In both cases, the tempo of steps has increased, the accordion more tightly closed, but the second embryonic phase of straightening is not the same.
Different neural dynamics are at work.
Working with Fabienne Lallouet, I studied an anencephalic human fetus without flexion of the base using CT scans, in order to verify whether its canals remained undifferentiated.41 A study undertaken in 2008 compared this fetus with normal Gorilla and Homo sapiens fetuses of the same craniofacial length at thirty-one weeks. The traditional expectation had been that simian traces would be in evident in human embryonic semi-circular canals. As it turned out, the conformation of these embryos failed to exhibit simian characteristics. This indicates that the morphogenesis of the canals follow a chronological and spatial cascade different from the apes. It is by means of this cascade that a delay from the cephalic pole is amplified all along the dorsal chord. The regulatory genes of hominid embryogenesis must have evolved from the axial embryonic patterns of the great apes.42 As a result, the caudal pole of the embryo forms later in Homo sapiens than in the great apes.
Going Straight
Neural straightening from Australopithecus to Homo is linked to the development of the nervous system. This explains the straightening of not only the base of the skull but also the whole vertebral column. The neural tube is formed by fibers lengthening along its path above the notochord. Complex movements at stage nineteen of embryonic development, corresponding to the axis of rotation, can be seen above the point at which the dorsal cord terminates. Hominization begins precisely at the cephalic limit of the dorsal cord.
Remarkable.
The sole vertebrate embryo in which the dorsal cord extremity is almost verticalized is that of Homo sapiens. This is a process that began around thirty-nine million years ago in an Asian species of prosimian that underwent a contraction in the base of its skull and a declination of its brain stem. This produced the first degree of neural straightening and cranio-facial contraction in the simians. Twenty-three million years ago, at least one African species of small gibbon-like simians underwent further contraction and declination. This produced the second degree of neural straightening. The embryonic dorsal cord was almost vertical among many species of great apes, remaining so until adulthood. This was presumably the case, at least, with respect to Australopithecus (4.5–1.977 mya).43 Thereafter the process accelerated, at an unprecedented rate. The lowered cerebellum and straightened brain stem is that of Homo sapiens, which Linnaeus named in 1758 and which emerged in East Africa 160,000 years ago. The evolutionary trajectory follows the straightening of the dorsal cord, but during the first stages of verticalization there was no dramatic accompanying increase in brain volume. Cranial volume is thus no longer the benchmark, or rubicon. The benchmark is, in fact, the straightening of the skull base.
The first stage is that of the hominin, which includes at least two embryogeneses, Australopithecus and Homo. The term is not Linnaean, but it avoids the contemporary confusion which includes gorillas and chimpanzees in the same family as Homo sapiens, yet excludes the orangutan. These groupings do not reflect observed morphogenetic structures and also serve to mask the presence of thresholds, namely, the discontinuities which have divided paleontologists. The trajectory of evolution from the first primates to Homo sapiens is defined by the increasing complexity of the nervous system. But this process was not gradual and not limited to the cerebral hemispheres. Neural embryogenesis increased in complexity, while the supporting tissues that would become the skeleton were transformed. Hence the thresholds and angular discontinuities. This is the process at the origin of neural straightening, in particular that of the cerebellum.
Embryonic axial straightening is a dynamic epiphenomenon resulting from a change in the gene regulation controlling the cephalocaudal axis. This evolution is not related to a change in habitat. Changes are produced in the amniotic fluid over a distance of several millimeters, reiterating a process that appeared in the genome with the emergence of the great apes. Australopithecus represents a threshold of neuronal complexity. The rotation of the neural tube was prolonged for longer than for any great ape, and thus the central nervous system became completely vertical. Permanent bipedalism appeared as a result. No paleontologist would suggest it was necessary for a species to become erect in order to begin walking exclusively on the ground. Gibbons and siamangs today are arboreal and walk easily on two legs.
An evolution similar to the emergence of australopithecines is seen with Telanthropus. This new conformation remains that of the Pithecanthropus, Sinanthropus, and Atlanthropus, but none of them attained the embryonic folding that belongs to Homo sapiens. Their basicranial axial skeleton is less verticalized, and this remains true of the Neanderthals (Eurasia, extinct around 28,000 years ago) and of Homo floresiensis (Indonesia, extinct around 80,000 years ago). Nothing in their neural and skeletal morphogenesis corresponds to the organizational structure of Homo sapiens.
It is necessary to name this ontogenesis. Do we call it Homo and exclude the embryogenesis of present humanity? Or do we give it a name like Paleoanthropus? I chose Homo because it is how, since 1964, all fossils that were once classified as Paleoanthropus are classified, regardless of cranial capacity and diet-related variations in tooth enamel.44
I have retained sapiens for the last threshold.
Considering the relevant parameters, it appears that Homo embryogenesis could not have been derived from australopithecine embryogenesis:
- The earliest angular values of the embryonic contraction (the accordion phase) of the Homo base are identical with those of australopithecines.45
- As can be seen in Paranthropus, further evolution of the australopithecine skull base occurred 1.8 million years ago (mya), separately, in eastern Africa (Paranthropus boisei) and South Africa (Paranthropus robustus), exhibiting a maximal embryonic accordion-like compression.46
- The Homo and sapiens embryonic thresholds are separated by at least 3 million years.
The modes and tempos of punctuated equilibria have accelerated between Australopithecus and Paranthropus (over 2 mya), on the one hand, and between Homo and Sapiens (over 3 mya), on the other. Because the earliest embryonic angular values are unchanged between Australopithecus and Homo, the latter could not have emerged from australopithecine embryogenesis.47
Misclassifying
Linnaean classification leads to gradualist and linear assumptions that have resulted in paleontologists looking for Homo characteristics in a species of Australopithecus without referring to the morphogenesis of its internal structure. For example, a skeleton from 3.6 mya was discovered in Ethiopia. Its anatomy was described in 2015 as close to that of Homo sapiens, but it has been kept within the genus Australopithecus. The base of the skull is missing, which would have allowed us to know from the semicircular canals whether it was of Homo embryogenesis or not.
Another example is an Ethiopian mandible from 2.8 mya, with a symphysis that is typical of the Australopithecus. Other of its features are considered to be adaptive traits related to masticatory function, and therefore to a diet similar to that of Homo. The fossil was thus attributed to the first Homo species, and the mandible said to be intermediate between the two genera.48 But how is it that the genus Homo could be applied to a lower jaw fragment of an Australopithecus adapted to an omnivorous diet without reference to the basicranial context and encephalization? In this case, function is prior to structure, but embryogenesis, which formed the structure, cannot be half-Australopithecus and half-Homo. It is one or the other, or that of another internally upright organism.
Mosaic gradualism has become a dogma that profoundly alters our understanding of the patterns and tempos of the process of humanization, instead of allowing us to begin by understanding the patterns of structural development before the structures begin to function, during morphogenesis.
Threshold Effects
When australopithecine embryogenesis emerged around 4 mya, the new organism was no longer a subspecies of great ape. It did not share a great ape’s neural embryogenesis, still visible today in gorillas, chimpanzees, and orangutans. The same is also true for Homo embryogenesis. There is no bone conformation that can be nested within previous embryonic organizations. The shapes of the structures are reorganized and innovations have appeared. This dynamic is typical of emergence in complex systems.
Sapiens embryogenesis is no longer that of any fossil Homo species. The axial skeleton reveals a new threshold of neural straightening and craniofacial contraction, with an increased complexity and reorganization of the central nervous system. The cerebellum is shifted even further toward the front and down. The occluso-postural equilibrium and the psychomotor development visible in, for example, Neanderthals, has also changed.
As Buffon and Cuvier observed, there is no unique pattern of anatomical organization among primates. The idea that there have been gradual transformations within the limits of a such an organization is absurd. This would mean that Homo sapiens has gone through all the strata of evolution—prosimian, ape, great ape, hominin. Homo sapiens is not a prosimian, nor is it a great ape. The conclusion that Homo sapiens is a primate indicates that prosimian innovations have been preserved. But it does not signify evolution within the limits of the same embryonic organization. Sapiens never passed through the stages of phylogenesis.
Linnaean classification is invisible in our embryogenesis.
If the word Homo must designate the paleoanthropes, then our embryogenesis is no longer Homo, but sapiens. This is true for each of the five great embryogeneses, or fundamental ontogeneses, since the emergence of Simian; each emerged from the preceding one, each time more complex in its neural organization. Numerous subspecies appear within the limits of the new axial embryogenesis, with different modes of locomotion and ways of life. Buffon, Cuvier, de Quatrefages, and Broca were right.
Conclusion
It is nothing short of remarkable that the ability to create second-degree stone tools emerged from the threshold of embryonic neural verticality. This is not an arbitrary boundary for distinguishing between Homo and other hominins, as is the case with the notion of a cerebral rubicon. The threshold is objective and allows for the deduction of a reorganization of the nervous system and its component neuronal networks with the sensors necessary for controlling the body’s equilibrium. In Homo sapiens, the connections between the cerebellar and cerebral neocortex are known, and it appears they participate in high-level cognitive functions, for example memory, dexterity, language, and reflection. Gestures such as walking and grasping become conscious with psychomotor development.
The great novelty here is the sudden change in posture of the cerebellum, and a new neuronal complexity; the cerebellum had to control its own balance. A new loop of complexity must have developed between the neocortex of the cerebellum and the brain. These connections could then have favored the development of new reflective cognitive capacities associated with movements, those of the hands in particular.
New manual chains of operation reflect a symbolic and conceptual level of thought attributed to the brain of the genus Homo. My suggestion is that the emergence of these capacities should be broadened to encompass the hominin stage, denoted by the verticalization of the cerebellum, such as for Australopithecus, Kenyanthropus, and Paranthropus. Although their brains were smaller than that of Homo habilis, they may have been capable of conceptual and creative innovations.49 Passing those first thresholds made possible the creative expression of ideas and concepts.
Crossing the final threshold, Sapiens, amplified those cognitive properties.
